
Dormansi benih dapat didefinisikan sebagai penghalang intrinsik untuk menyelesaikan perkecambahan benih walaupun benih dalam kondisi yang mendukung perkecambahan (misalnya suhu, kelembaban, cahaya sesuai). Dormansi benih mengontrol waktu perkecambahan sebagai respons terhadap kondisi dan perubahan musim/iklim serta memainkan peran penting dalam evolusi benih tanaman. Waktu perkecambahan sangat mempengaruhi laju dimana spesies dapat memperluas jangkauan/penyebaran mereka dan memainkan peran penting dalam kelangsungan hidup atau kepunahan selama perubahan iklim (Donohue 2005). Penelitian Baskin dan Baskin (2004), telah mengusulkan sistem klasifikasi ekofisiologi komprehensif untuk dormansi benih yang meliputi lima kelas: fisiologis (PD), morfologis (MD), morfofisiologis (MPD), fisik (PY) dan kombinasional (PY+PD).
Dormansi morfologis (MD) dapat terlihat jelas pada benih dengan ukuran embrio yang relatif kecil dibandingkan dengan ukuran seluruh benih (Gambar 1). Hal ini ditunjukkan dari rasio embrio terhadap biji (Rasio E:S) yang menggambarkan ukuran relatif embrio di dalam benih. Rasio E:S yang tinggi (misalnya 0,9) menunjukkan embrio mengisi sebagian besar volume benih, sedangkan rasio E : S yang rendah (misalnya 0,1) menunjukkan embrio kecil dan jaringan penyimpanan nutrisi mengisi sebagian besar volume benih (Gambar 1). Pada Angiospermae jaringan nutrisi terdapat pada perisperm dan endosperm, sedangkan Gymnospermae terdapat pada megagametofit. Benih dengan rasio E:S yang rendah seringkali memiliki waktu perkecambahan yang lama (>1 bulan).
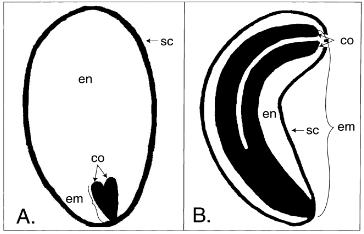
Gambar 1 Contoh perbedaan ukuran embrio dengan ukuran keseluruhan benih pada biji (A) Thalictrum polygamum (Ranunculaceae). (B) Ruta graveolens (Rutaceae). EM (embrio); EN (endosperma); SC (kulit biji); CO (kotiledon) (Sumber: Forbis et al. 2002).
Angiospermae dan Gymnospermae mengalami tren evolusi peningkatan ukuran embrio. Dalam penelitian Forbis et al. (2002), dilakukan rekonstruksi keadaan leluhur dari karakter kontinu 179 famili (dari 1222 spesies angiospermae yang masih hidup) dengan menggunakan Rasio E:S. Hasil analisis mereka (Gambar 2) menunjukkan bahwa Rasio E:S meningkat pada keturunan angiospermae dibandingkan dengan nenek moyang angiospermae. Fenomena ini juga terlihat pada gymnospermae (Gambar 3), nenek moyangnya memiliki rasio ukuran embrio yang kecil saat awal penyebaran kemudian mengalami peningkatan ukuran embrio di keturunannya (Conifers, Cycads, Ginkgos, Gnetales dll.). Cycad dan ginkgo, memiliki embrio yang lebih kecil daripada beberapa taksa gymnospermae lain, seperti Callitropsis, Picea, Pinus dan Juniperus. Dari keseluruhan informasi tersebut dapat disimpulkan bahwa dormansi morfologi adalah tipe dormansi leluhur di antara gymnospermae dan angiospermae. Evolusi lebih besar ukuran embrio kemungkinan besar mengakibatkan terjadinya non-dorman pada biji, sehingga embrio tidak perlu tumbuh sebelum berkecambah. Peningkatan ukuran relatif embrio juga menjadi salah satunya penentu utama (atau persyaratan) untuk evolusi kelas dormansi benih lainnya (Finch-Savage dan Leubner Metzger 2006).
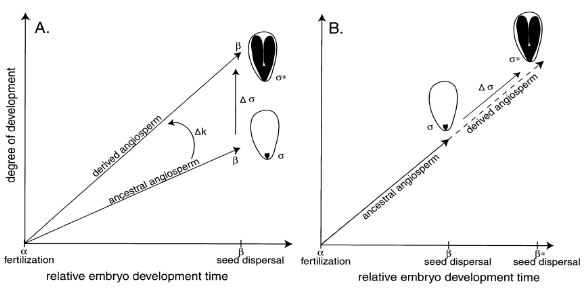 Gambar 2 Dua skenario alternatif untuk perubahan perkembangan morfologi embrio antara spesies angiospermae leluhur hipotesis dan spesies angiospermae turunan hipotesis. A) permulaan perkembangan (pembuahan); B) perimbangan perkembangan (penyebaran biji) (Sumber: Forbis et al. 2002).
Gambar 2 Dua skenario alternatif untuk perubahan perkembangan morfologi embrio antara spesies angiospermae leluhur hipotesis dan spesies angiospermae turunan hipotesis. A) permulaan perkembangan (pembuahan); B) perimbangan perkembangan (penyebaran biji) (Sumber: Forbis et al. 2002).
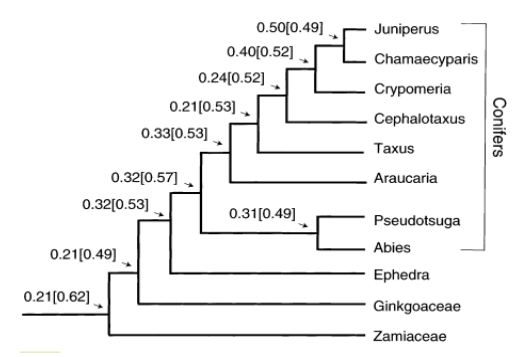 Gambar 3 Rekonstruksi nenek moyang gymnospermae bersamaan dengan nilai kuadrat terkecil (GSL) yang menyatakan rasio embrio dan biji. Analisis dilakukan pada semua cabang dengan angka-angka pada simpul mewakili nilai E:S dan angka dalam tanda kurung mewakili nilai kesalahan standar (Sumber: Forbis et al. 2002).
Gambar 3 Rekonstruksi nenek moyang gymnospermae bersamaan dengan nilai kuadrat terkecil (GSL) yang menyatakan rasio embrio dan biji. Analisis dilakukan pada semua cabang dengan angka-angka pada simpul mewakili nilai E:S dan angka dalam tanda kurung mewakili nilai kesalahan standar (Sumber: Forbis et al. 2002).
Dormansi Fisiologis (PD) adalah jenis dormansi yang paling melimpah dan ditemukan pada benih semua kelompok gymnospermae dan angiospermae (Gambar 4). PD dibagi menjadi beberapa jenis dengan bentuk yang paling umum pada angiospermae dan gymnospermae adalah PD nondeep. PD nondeep terjadi karena faktor fisiologis pada embrio dan/atau mantel benih (Holdsworth et al. 2008). Dormansi mantel benih dimediasi oleh salah satu lapisan penutup (endosperma dan ⁄ atau testa). Asam absisat (ABA) mengatur PD nondeep yang dimediasi oleh mantel pada benih baik gymnospermae maupun angiospermae (Kucera et al. 2005). Gymnospermae dan angiospermae memiliki kesamaan mekanisme molekuler terkait ABA. Faktor transkripsi yang berhubungan dengan ABA (ABA13/VP1) tersebar luas di antara tanaman hijau dan terlibat dalam mengatur dormansi benih dan tunas angiosperma dan gymnospermae. Ketergantungan ABA adalah sifat plesiomorfik untuk angiospermae dan gymnospermae (Graeber et al. 2009). Embrio yang dipotong dari biji PD nondeep akan berkecambah secara normal dan perlakuan dengan giberelin (GA) juga akan mematahkan dormansi. Selain itu, tergantung pada spesiesnya, dormansi dapat dipatahkan dengan skarifikasi (pengikisan atau pemotongan lapisan penutup), pematangan (periode penyimpanan kering udara), stratifikasi hangat atau dingin.
Daftar Pustaka
Baskin JM, Baskin CC. 2004. A classification system for seed dormancy. Seed Science Research. 14: 1-16.
Donohue K. 2005. Niche construction through phenological plasticity: life history dynamics and ecological consequences. New Phytologist. 166: 83-92.
Finch-Savage WE, Leubner-Metzger G. 2006. Seed dormancy and the control of germination. New Phytologist. 171: 501-523.
Forbis TA, Floyd SK, deQueiroz A. 2002. The evolution of embryo size in angiosperms and other seed plants: implications for the evolution of seed dormancy. Evolution. 56: 2112-2125.
Graeber K, Linkies A, Muller K, Wunchova A, Rott A, Leubner-Metzger G. 2010. Cross-species approaches to seed dormancy and germination: conservation and biodiversity of ABA-regulated mechanisms and the Brassicaceae DOG1 genes. Plant Molecular Biology. 73: 67-87.
Holdsworth MJ, Bentsink L, Soppe WJJ. 2008. Molecular networks regulating Arabidopsis seed maturation, after-ripening, dormancy and germination. New Phytologist. 179: 33-54.
Kucera B, Cohn MA, Leubner-Metzger G. 2005. Plant hormone interactions during seed dormancy release and germination. Seed Science Research. 15: 281-307.
Linkies A, Graeber K, Knight C, Leubner‐Metzger G. 2010. The evolution of seeds. New Phytologist. 186(4): 817-831.
Created by: Seedling Group




